科研进展
噬菌体协助细菌帝国之间建立“界限”
发布时间:2019-05-27
作者:LMB
细菌是“高情商”的生物,懂得“社交”。他们以团结协作的群体形式生活在自然环境中,以抵抗来自外界环境的压力。同一细菌群体中还存在自我识别能力,可以区分哪些是同一家族,同时共同抵抗“异族”。具有扩散能力的化学信号分子和细菌的表面受体可作为细菌群体识别的语言和工具。
噬菌体是地球上数量最多的一种细菌病毒。噬菌体的生存依附于细菌,但它的繁殖和扩散又依赖于细菌的死亡。噬菌体与细菌永不停息的相爱相杀共同谱写了生物进化的篇章,而广泛使用的限制性内切酶和CRISPR系统是二者相爱相杀的最好证据。
日前,中科院南海海洋所的王晓雪课题组和宾夕法尼亚州立大学的Thomas Wood 课题组在4月16日的Cell Reports杂志上报道了细菌游动过程中使用感染它们的病毒 (即噬菌体)进行自我识别的新方式。这是细胞使用病毒将自己与近源的竞争对手区分开来的第一个证据。
研究的想法开始于王晓雪等准备运动平板,科研人员发现当把不同大肠杆菌 K-12 BW25113菌株接种在同一运动平板上时,经过一段时间,菌株在相遇的区域建立起一道“防御线”,形成了一道类似于“国界”的分界线(图1,红色箭头)。
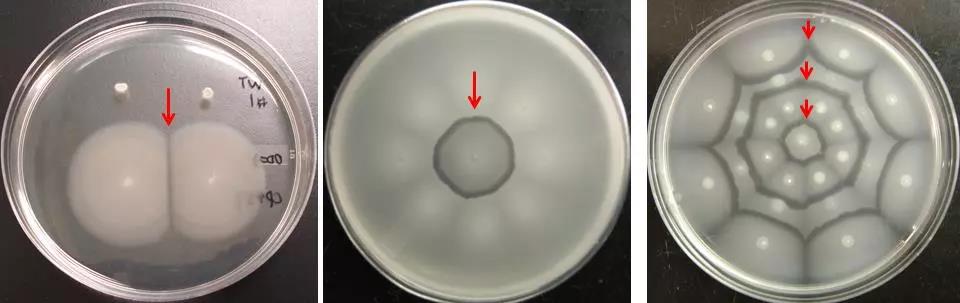
图1 大肠杆菌自我识别的“分界线”(红色箭头)
为了揭示分界线形成的机制,科研人员对已有的大肠杆菌 K-12 突变文库的4296个单基因敲除的菌株进行筛选。幸运的是,他们发现当细菌中携带的突变影响到噬菌体复制所需的基因,分界就完全消失了。研究人员推测,“分界线”可能是通过烈性噬菌体引起的细胞裂解形成的。
对边界线处的群体进行研究发现,在长期的进化过程,一个类似于T1的噬菌体(重新命名为SW1)被细菌捕获,将其基因组整合到了细菌基因组上。在运动平板上,当携带SW1噬菌体的大肠杆菌与不携带SW1噬菌体的大肠杆菌相遇时,一部分细菌中SW1噬菌体会重新被激活,重新释放到环境中,去攻击不带有此噬菌体的细胞(外族,没有标记“SW1”)并杀死这些细胞,从而在“交战区”形成了没有活细菌而存在大量活跃的SW1的透明区,也就是平板上看到的分界线(demarcation line)(图2A)。
分界线一旦形成,界内的细菌可以在这道屏障的保护下进行繁殖和搜寻营养物质,互不干扰。而携带有SW1噬菌体的细菌可以帮助细菌抵御其它噬菌体的入侵(图2B),并在特定的情况下提供争夺营养的优势条件。
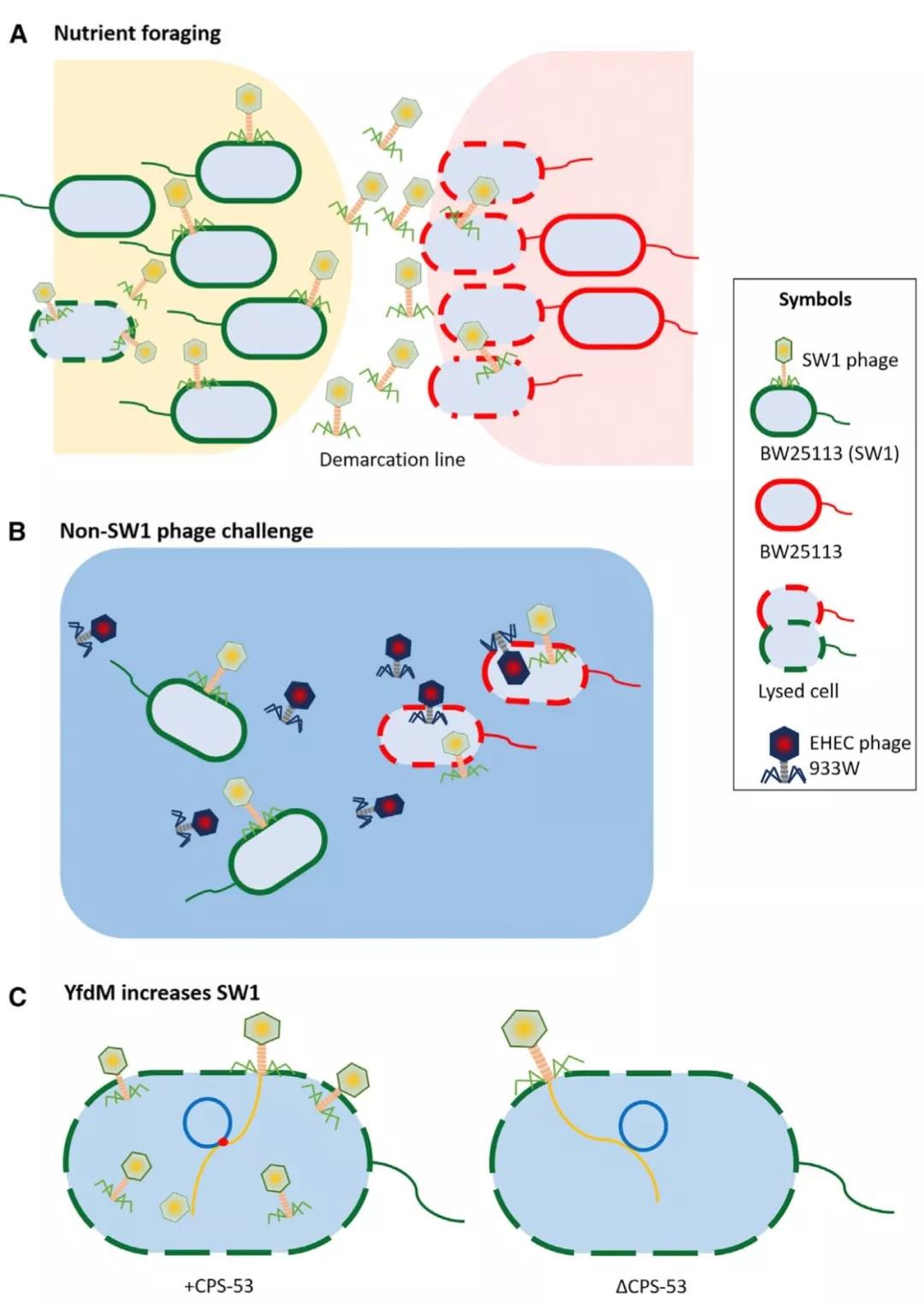
图2 SW1病毒对大肠杆菌竞争优势的贡献。(A)游动过程中,无SW1病毒的细菌与携带SW1的细菌相遇,被对方释放出的SW1裂解,从而形成分界线;(B)携带SW1病毒在其他噬菌体入侵时为细菌提供保护;(C)原噬菌体CPS53的YfdM促进SW1的繁殖。
利用王晓雪等前期构建的大肠杆菌K-12中9个隐匿型噬菌体的缺失突变株(Wang et al., 2010 Nature Communications)进行检测时,研究人员发现其中1个古老的隐匿型噬菌体CPS-53携带的YfdM蛋白,是负责细菌自我识别的关键蛋白。他们发现,将携带SW1的细菌细胞暴露在 YfdM 中,会刺激噬菌体颗粒的产生,并导致缺乏 SW1细胞的裂解。相反的,较高浓度的 YfdM 或噬菌体颗粒会在细菌细胞之间产生更为明显的分界线(图2C)。
研究结果表明,SW1 通过使用宿主中的CPS-53神秘防御蛋白YfdM 进行控制“分界线”的形成。而神秘的防御蛋白来自于一个更加古老的噬菌体。
这种新的自我识别机制允许细菌群体与“亲属”进行交流,同时对抗“外族”,这种交流在对营养需求、毒力表达、密度感应和生物膜形成中可能非常重要。细菌能够通过化学信号的交流将自己与其他细菌区分开来,这项新的研究展示了细胞利用病毒来区分自己和周围环境中密切相关的细菌。因此,应该重新评估病毒与其细胞宿主之间的关系,因为病毒感染可能在某些竞争条件下对宿主更为有利。
在未来的研究中,研究人员计划研究SW1如何避免攻击其宿主细胞,而不杀死缺乏病毒的细菌。由于人和动物肠道中定殖的细菌多可在肠道表明游动,为了获得更多的生存空间,它们将自己携带的噬菌体作为武器,攻击其它近源细菌,系统的研究将有助于了解细胞如何竞争或许可以更好地利用病毒来对抗细菌感染,对肠道致病菌的治疗具有一定的参考价值。
论文题目Phages Mediate Bacterial Self-Recognition。文章链接:https://www.cell.com/cell-reports/fulltext/S2211-1247(19)30400-0
附件下载: